Какие продукты образуются при полном гидролизе нуклеиновых кислот
Полный гидролиз - нуклеиновая кислота
Полный гидролиз - нуклеиновая кислота
Cтраница 1
Полный гидролиз нуклеиновых кислот приводит к образованию эквивалентных количеств фосфорной кислоты, пентозы и пу-риновых или пиримидиновых оснований. При гидролизе нуклеиновых кислот ферментами желудочного или поджелудочного сока образуются мононуклеотиды. [1]
Продуктами полного гидролиза нуклеиновых кислот являются гетероциклические азотистые основания, углеводы ( рибоза или дезоксирибоза) и фосфорная кислота. Следовательно, остатки этих соединений являются структурными компонентами нуклеиновых кислот. [2]
При полном гидролизе нуклеиновых кислот образуются фосфорная кислота, сахар, ппрпмидины и пуриновые основания. Сахар - входящий в состав нуклеиновых кислот цитоплазмы, представляет собой D-рибозу; его содержат также нуклеиновые кислоты, полученные из дрожжей. Эти нуклеиновые кислоты называют рибонуклеиновыми кислотами. [3]
При полном гидролизе нуклеиновых кислот образуются фосфорная кислота, сахар, пиримидины и пуриновые основания. Сахар, входящий в состав нуклеиновых кислот цитоплазмы, представляет собой D-рибозу; его содержат также нуклеиновые кислоты, полученные из дрожжей. Эти нуклеиновые кислоты называют рибонуклеиновыми кислотами. [4]
В результате полного гидролиза нуклеиновых кислот минеральными кислотами образуется смесь пиримидинов и пуринов ( см. ниже), сахар ( а именно пентоза) и фосфорная кислота. Таким образом, нуклеиновые кислоты построены как продукты поликонденсации более простых компонентов, называемых нуклеотидами. Последние получаются при мягком гидролизе нуклеиновых кислот разбавленным аммиаком. Следовательно, нуклеиновые кислоты являются полипуклеотидами. [5]
Таким образом, при полном гидролизе нуклеиновой кислоты получается смесь, содержащая фосфорную кислоту, альдопентозу и гетероциклические основания. [6]
Какие продукты образуются при: а) частичном; б) полном гидролизе нуклеиновых кислот. [7]
Нуклеиновые кислоты - высокомолекулярные соединения ( мол. При полном гидролизе нуклеиновых кислот образуется смесь из азотсодержащих гетероциклических оснований ( пиримидиновых и пуриновых оснований, см. § 170), моносахаридов - пентоз ( рибоза или дез-оксирибоза) и фосфорной кислоты. [8]
При полном гидролизе нуклеиновых кислот образуется смесь из азотсодержащих гетероциклических оснований ( пиримидиновых и пуриновых оснований), моносахаридов - пентоз ( рибоза или дезоксирибоза) и фосфорной кислоты. [9]
При полном гидролизе нуклеиновых кислот образуется смесь окси-и аминопроизводных пиримидина и пурина ( пиримндиновые и пуриновые основания), моносахарид - пентоза ( рибоза или дезок-сирибоза) и фосфорная кислота. [10]
При полном гидролизе нуклеиновых кислот образуются смесь окси - и аминопроизводных пиримидина и пурина ( пиримидиновые и пуриновые основания), моносахарид - пентоза ( рибоза или дезоксирибоза) и фосфорная кислота. [11]
При полном гидролизе нуклеиновых кислот образуется смесь окси-и аминопроизводных пиримидина и пурина ( пиримидиновые и пуриновые основания), моносахарид - пентоза ( рибоза или дезок-сирибоза) и фосфорная кислота. [12]
Прежде чем перейти к изложению вопроса о структуре нуклеиновых кислот, следует рассмотреть строение отдельных соединений, входящих в состав молекулы нуклеиновой кислоты. В результате полного гидролиза нуклеиновых кислот образуются пири-мидиновые и пуриновые основания, сахар и фосфорная кислота. Частичный гидролиз приводит к образованию нуклеозидов и нуклеотидов. Все эти соединения рассмотрены ниже. [13]
Нуклеозиды в условиях кислотного гидролиза распадаются на азотистые основания и сахара. Таким образом, при полном гидролизе нуклеиновых кислот образуются азотистые основания, моносахарид пентоза ( рибоза или дезоксирибоза) и фосфорная кислота. [14]
В слизистой кишечника содержится активный фермент 5 -рибонуклеотид-фосфогидролаза ( 5 -нуклеотидаза), гидролизующая 5 -рибонуклеотиды на нуклеозиды и фосфорную кислоту. Под действием 5 -рибонуклеотид-фосфогидролазы происходит, по-видимому, гидролиз 5 -рибонуклеотидов на поверхности слизистой оболочки кишечника. Образующиеся при этом нуклеозиды легко всасываются через клеточные мембраны. В клетках кишечных стенок нуклеозиды расщепляются до пентоз и пуриновых или пиримидино-вых оснований. Продукты полного гидролиза нуклеиновых кислот поступают с кровью в печень и другие ткани организма, где происходит биосинтез новых нуклеиновых кислот, а также дальнейшие процессы расщепления. В результате промежуточного обмена в организме человека пуриновые основания превращаются в мочевую кислоту, а пиримиди-новые основания дают СО2, NHi, р-аланин и ( З - аминоизомасляную кислоту. [15]
Страницы: 1 2
нуклеиновых кислот | Определение, функции, структура и типы
Нуклеиновая кислота , химическое соединение природного происхождения, способное расщепляться с образованием фосфорной кислоты, сахаров и смеси органических оснований (пурины и пиримидины). Нуклеиновые кислоты являются основными молекулами клетки, несущими информацию, и, управляя процессом синтеза белка, они определяют унаследованные характеристики каждого живого существа. Двумя основными классами нуклеиновых кислот являются дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновая кислота (РНК).ДНК - это главный план жизни и генетический материал всех свободноживущих организмов и большинства вирусов. РНК - это генетический материал некоторых вирусов, но она также обнаружена во всех живых клетках, где играет важную роль в определенных процессах, таких как создание белков.
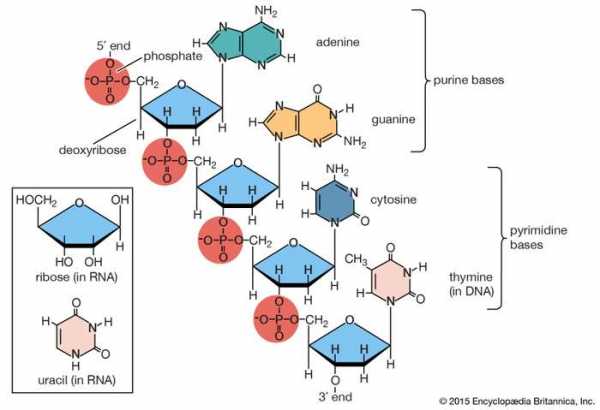 полинуклеотидная цепь дезоксирибонуклеиновой кислоты (ДНК) Часть полинуклеотидной цепи дезоксирибонуклеиновой кислоты (ДНК). На вставке показаны соответствующие пентозный сахар и пиримидиновое основание в рибонуклеиновой кислоте (РНК). Encyclopædia Britannica, Inc.
полинуклеотидная цепь дезоксирибонуклеиновой кислоты (ДНК) Часть полинуклеотидной цепи дезоксирибонуклеиновой кислоты (ДНК). На вставке показаны соответствующие пентозный сахар и пиримидиновое основание в рибонуклеиновой кислоте (РНК). Encyclopædia Britannica, Inc. Популярные вопросы
Что такое нуклеиновые кислоты?
Нуклеиновые кислоты - это встречающиеся в природе химические соединения, которые служат основными молекулами, несущими информацию в клетках. Они играют особенно важную роль в управлении синтезом белка. Двумя основными классами нуклеиновых кислот являются дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновая кислота (РНК).
Какова основная структура нуклеиновой кислоты?
Нуклеиновые кислоты - это длинные цепочечные молекулы, состоящие из ряда почти идентичных строительных блоков, называемых нуклеотидами.Каждый нуклеотид состоит из азотсодержащего ароматического основания, присоединенного к пентозному (пятиуглеродному) сахару, которое, в свою очередь, присоединено к фосфатной группе.
Какие азотсодержащие основания встречаются в нуклеиновых кислотах?
Каждая нуклеиновая кислота содержит четыре из пяти возможных азотсодержащих оснований: аденин (A), гуанин (G), цитозин (C), тимин (T) и урацил (U). A и G относятся к пуринам, а C, T и U называются пиримидинами. Все нуклеиновые кислоты содержат основания A, C и G; Однако T находится только в ДНК, а U - в РНК.
Когда были открыты нуклеиновые кислоты?
В этой статье рассматривается химия нуклеиновых кислот, описываются структуры и свойства, которые позволяют им служить передатчиками генетической информации. Для обсуждения генетического кода см. Наследственность , а для обсуждения роли нуклеиновых кислот в синтезе белка см. Метаболизм .
Нуклеотиды: строительные блоки нуклеиновых кислот
Основная структура
Нуклеиновые кислоты - это полинуклеотиды, то есть длинные цепочечные молекулы, состоящие из ряда почти идентичных строительных блоков, называемых нуклеотидами.Каждый нуклеотид состоит из азотсодержащего ароматического основания, присоединенного к пентозному (пятиуглеродному) сахару, которое, в свою очередь, присоединено к фосфатной группе. Каждая нуклеиновая кислота содержит четыре из пяти возможных азотсодержащих оснований: аденин (A), гуанин (G), цитозин (C), тимин (T) и урацил (U). A и G классифицируются как пурины, а C, T и U вместе называются пиримидинами. Все нуклеиновые кислоты содержат основания A, C и G; Однако T находится только в ДНК, а U - в РНК. Сахар-пентоза в ДНК (2'-дезоксирибоза) отличается от сахара в РНК (рибоза) отсутствием гидроксильной группы (OH) на 2'-атоме углерода сахарного кольца.Без присоединенной фосфатной группы сахар, присоединенный к одному из оснований, известен как нуклеозид. Фосфатная группа соединяет последовательные остатки сахара, соединяя 5'-гидроксильную группу одного сахара с 3'-гидроксильной группой следующего сахара в цепи. Эти нуклеозидные связи называются фосфодиэфирными связями и одинаковы в РНК и ДНК.
Нуклеотиды синтезируются из легкодоступных предшественников в клетке. Рибозофосфатная часть пуриновых и пиримидиновых нуклеотидов синтезируется из глюкозы через пентозофосфатный путь.Сначала синтезируется пиримидиновое кольцо из шести атомов, которое затем присоединяется к рибозофосфату. Два кольца в пуринах синтезируются, будучи присоединенными к фосфату рибозы во время сборки нуклеозидов аденина или гуанина. В обоих случаях конечный продукт представляет собой нуклеотид, несущий фосфат, связанный с 5'-атомом углерода на сахаре. Наконец, специальный фермент, называемый киназой, добавляет две фосфатные группы, используя аденозинтрифосфат (АТФ) в качестве донора фосфата, с образованием рибонуклеозидтрифосфата, непосредственного предшественника РНК.В случае ДНК 2'-гидроксильная группа удаляется из рибонуклеозиддифосфата с образованием дезоксирибонуклеозиддифосфата. Дополнительная фосфатная группа из АТФ затем добавляется другой киназой с образованием дезоксирибонуклеозидтрифосфата, непосредственного предшественника ДНК.
Получите эксклюзивный доступ к контенту из нашего первого издания 1768 с вашей подпиской. Подпишитесь сегодняВо время нормального клеточного метаболизма РНК постоянно производится и разрушается. Остатки пурина и пиримидина повторно используются несколькими путями восстановления для создания большего количества генетического материала.Пурин спасается в форме соответствующего нуклеотида, тогда как пиримидин спасается как нуклеозид.
.Глава 5. Углеводы 1
Глава 5. Углеводы 1
1. ВВЕДЕНИЕ
2. КЛАССИФИКАЦИЯ И ХИМИЯ
3. УГЛЕВОДНЫЙ МЕТАБОЛИЗМ В РЫБЕ
4. ССЫЛКИ
K. W. Chow
Продовольственная и сельскохозяйственная организация
Рим, ИталияДж. Э. Халвер
Вашингтонский университет
Сиэтл, Вашингтон
1/ Лекцию прочитал Дж.Э. Халвер
Углеводы представляют собой широкую группу веществ, в которую входят сахара, крахмалы, камеди и целлюлозы. Общие свойства углеводов заключаются в том, что они содержат только элементы углерод, водород и кислород, и что при их сгорании образуется двуокись углерода плюс одна или несколько молекул воды.
Самыми простыми углеводами являются трехуглеродные сахара, которые играют важную роль в промежуточном метаболизме, а самыми сложными являются встречающиеся в природе полисахариды, в основном растительного происхождения.В рационе животных и рыбы значимы два класса полисахаридов:
(а) структурные полисахариды, которые усваиваются травоядными видами - целлюлоза, лигнин, декстраны, маннаны, инулин, пентозаны, пектиновые кислоты, альгиновые кислоты, агар и хитин; а также(б) универсально усваиваемые полисахариды - в основном крахмал.
Углеводы составляют три четверти биомассы растений, но присутствуют в организме животных лишь в небольших количествах в виде гликогена, сахаров и их производных.Гликоген часто называют животным крахмалом, поскольку он отсутствует в растениях. Производные моносахариды, такие как сахарные кислоты, аминосахары и дезоксисахары, являются составными частями всех живых организмов.
2.1 Пентозы
2.2 Гексозы
2.3 Дисахариды
2.4 Олигосахариды
2.5 Полисахариды
Углеводы обычно классифицируются по степени сложности. Следовательно, свободные сахара, такие как глюкоза и фруктоза, называются моносахаридами; сахароза и мальтоза, дисахариды; а также крахмалы и целлюлозы, полисахариды.Углеводы с короткими цепями, такие как рафиноза, стахиоза и вербаскоза, которые представляют собой три, четыре и пять полимеров сахаров соответственно, классифицируются как олигосахариды.
2,1 Пентозы
Пентозы - это пятиуглеродные сахара, редко встречающиеся в природе в свободном состоянии. У растений они встречаются в полимерных формах и все вместе известны как пентозаны. Таким образом, ксилоза и арабиноза входят в состав пентозанов, присутствующих в растительных волокнах и растительных камедях, соответственно. Поскольку сахарные фрагменты в нуклеиновых кислотах и рибофлавине, рибоза и дезоксирибоза являются незаменимыми составляющими жизненного процесса.D-рибоза имеет следующую химическую структуру:
|
|
| D-рибоза |
2,2 Гексозы
Гексозы составляют большую группу сахаров. Основные из них: глюкоза, фруктоза, галактоза и манноза. В то время как глюкоза и фруктоза в природе свободны, галактоза и манноза встречаются только в комбинированной форме. Гексозы делятся на альдозы и кетозы в зависимости от того, имеют ли они альдегидные или кетоновые группы.Таким образом, глюкоза представляет собой альдо-сахар, а фруктоза - кето-сахар. Присутствие асимметричных центров во всех сахарах с тремя или более атомами углерода приводит к стереоизомерам. Галактоза и манноза являются стереоизомерами глюкозы, которая теоретически является только одним из 16 стереоизомеров. Поскольку кетогексозы имеют только три асимметричных центра, фруктоза является одним из восьми стереоизомеров. Химические конфигурации четырех упомянутых гексоз следующие:
|
|
|
|
|
| D-глюкоза | D-галактоза | D-манноза | D-фруктоза |
Общее явление, известное как мутаротация, наблюдается в различных пентозах и гексозах, а также в некоторых дисахаридах.Например, было установлено, что существуют два изомера D-глюкозы, следовательно, требуется дополнительный асимметричный центр в этом сахаре. Стало очевидно, что D-глюкоза и большинство других сахаров имеют циклическую структуру. Положение гидроксильной группы по отношению к кольцевому кислороду характеризует эту дополнительную конфигурацию модификации. По соглашению, расположение гидроксильной группы на атоме углерода 1 на той же стороне структуры, что и кислородное кольцо, описывает -модификацию; и расположение той же гидроксильной группы на противоположной стороне кольцевого кислорода описывает b-модификацию.
|
|
|
| a -D-глюкоза | b -D-глюкоза |
Карбогидразы, которые катализируют гидролиз гликозидных связей простых гликозидов, олигосахаридов и полисахаридов, часто проявляют специфичность в отношении конфигурации субстрата. Как мы увидим позже, специфичность ферментативного гидролиза некоторых олигосахаридов помогает объяснить плохое использование этого класса углеводов в питании рыб.
Сахара, содержащие альдо- или кетогруппу, способны восстанавливать медь в щелочных растворах (раствор Фелинга) с образованием кирпично-красного цвета ионов одновалентной меди. Эти сахара называются восстанавливающими сахарами, и реакция, хотя и не специфична для восстанавливающих сахаров, может использоваться как для качественного, так и для количественного определения.
Глюкоза широко распространена в небольших количествах во фруктах, растительных соках и меде. Он коммерчески производится путем кислотного или ферментативного гидролиза зерновых и корневых крахмалов.Глюкоза представляет особый интерес для питания, потому что это конечный продукт переваривания углеводов у всех нежвачных животных, включая рыбу.
Фруктоза - единственная важная кетогексоза, которая в свободном состоянии находится вместе с глюкозой в созревающих фруктах и меде. В сочетании с глюкозой образует сахарозу. Фруктоза несколько слаще, чем сахароза, и ее в промышленных масштабах производят в возрастающих количествах в качестве подсластителя.
Галактоза содержится в молоке в сочетании с глюкозой.Он также присутствует в олигосахаридах растительного происхождения в сочетании с глюкозой и фруктозой.
Манноза присутствует в некоторых полисахаридах растений, которые собирательно называются маннанами.
2.3 Дисахариды
Дисахариды - это продукты конденсации двух молекул моносахаридов. Сахароза - преобладающий дисахарид, встречающийся в свободной форме, и основное вещество сахарного тростника и сахарной свеклы. Также он образуется при прорастании семян бобовых.Другие распространенные дисахариды - мальтоза и лактоза. Мальтоза представляет собой димер глюкозы, а лактоза - сополимер галактозы и глюкозы. Две молекулы глюкозы в мальтозе удерживаются вместе в гликозидной связи a -1,4, тогда как две гексозные составляющие галактозы связаны в положении b -1,4. В сахарозе глюкоза и фруктоза объединены в связь -1,2. Сокращенное название сахарозы - D-Glu- (a, 1® 2) -D-Fru.
a -Мальтоза
b -Лактоза
Сахароза
2.4 Олигосахариды
Олигосахариды рафиноза, стахиоза и вербаскоза присутствуют в значительных количествах в семенах бобовых. Рафиноза, которая является наиболее распространенной из трех, состоит из одной молекулы глюкозы, связанной с молекулой сахарозы в положении a -1,6. Его сокращенное химическое название - a -D-Gal (1® 6) -a - D -Glu - (1® 2) - b -D-Fru. Дальнейшее удлинение цепи на конце галактозы с другой молекулой галактозы приведет к образованию стахиозы. Все эти связи галактоза-галактоза находятся в положении a-l, 6, и переваривание этих олигосахаридов животными требует высокоспецифического фермента, вырабатываемого не самими животными, а некоторыми бактериями, присутствующими в кишечнике животных.Постепенное исчезновение олигосахаридов из котелидонов семян бобовых во время прорастания является частью сложного процесса, начинающегося с поглощения воды семенами. Это поглощение влаги высвобождает гибберелловую кислоту, которая, в свою очередь, активирует ДНК в семенах, тем самым запуская жизненный цикл растения. ДНК направляет производство -галактозидазы, которая необходима для гидролиза этих олигосахаридов. Любое вмешательство в процесс транскрипции ДНК блокирует производство ферментов и будет подтверждено продолжающимся старением семян и сохранением олигосахаридов в семенных котелидонах.
2,5 Полисахариды
Полисахариды представляют собой большую группу сложных углеводов, которые являются продуктами конденсации неопределенного числа молекул сахара. Различные подгруппы довольно плохо определены, и нет согласия по их классификации. Большинство полисахаридов нерастворимы в воде. При гидролизе кислотами или ферментами они в конечном итоге дают составляющие их моносахариды.
Крахмал представляет собой высокомолекулярный полимер D-глюкозы и является основным резервным углеводом в растениях.Большинство крахмалов состоят из смеси двух типов полимеров, а именно; амилоза и амилопектин. Пропорция амилозы и амилопектина обычно составляет одну часть амилозы и три части амилопектина. Ферменты, способные катализировать гидролиз крахмала , присутствуют в пищеварительном секрете животных и рыб внутри их клеток. Α-амилазы, которые обнаруживаются практически во всех живых клетках, случайным образом расщепляют связи a -D- (1® 4) и в конечном итоге вызывают полное превращение молекулы крахмала в восстанавливающие сахара.Основные α-амилазы животного происхождения - это те, которые вырабатываются слюнной железой и поджелудочной железой. Крахмал нерастворим в воде и окрашивается йодом в синий цвет.
Гликоген - единственный сложный углевод животного происхождения. Он присутствует в ограниченных количествах в печени и мышцах и действует как легкодоступный источник энергии.
Декстрины представляют собой промежуточные соединения, образующиеся в результате неполного гидролиза или переваривания крахмала. Присутствие -D- (1® 6) связей в амилопектине и неспособность -амилазы расщеплять эти связи приводят к образованию низкомолекулярных углеводных сегментов, называемых предельными декстринами.На эти остатки действуют прежде всего ацидофильные бактерии пищеварительного тракта.
Целлюлоза состоит из длинных цепей глюкозных единиц, удерживаемых вместе связями b -D- (1® 4). Ферменты, которые расщепляют эти связи, обычно не присутствуют в пищеварительном секрете животных и рыб, хотя считается, что некоторые виды моллюсков вырабатывают целлюлазу, фермент, катализирующий гидролиз целлюлозы. Микроорганизмы, продуцирующие целлюлазу, присутствующие в кишечнике травоядных животных и рыб, придают своим животным-хозяевам способность использовать в качестве пищи трудноусвояемую целлюлозу.
Другими широко распространенными сложными полисахаридами являются гемицеллюлозы и пентозаны. Гемицеллюлоза представляет собой группу углеводов, включая арабан, ксилан, некоторые гексозаны и полиурониды. Эти вещества, как правило, менее устойчивы к химической обработке и подвергаются некоторой степени ферментативному гидролизу во время нормальных пищеварительных процессов. Пентозаны представляют собой полимеры ксилозы или арабинозы в составе структурного материала растений и растительных камедей соответственно.
3.1 Пищеварение, абсорбция и хранение
3.2 Прочие факторы Влияние на метаболизм
3.3 Преобразование энергии
Большая часть углеводов, которые входят в рацион животных, включая рыбу, имеют растительное происхождение. Следовательно, хищные рыбы, такие как атлантический лосось и желтохвост японская, имеют мало углеводов. Действительно, эксперименты показали, что эти виды плохо приспособлены для обработки значительных количеств сырых углеводов в своем рационе.С другой стороны, всеядные животные, такие как карп и канальный сом, способны переваривать изрядное количество углеводов в своем рационе. Белый амур, травоядное животное, питается в основном вегетарианской диетой.
3.1 Пищеварение, абсорбция и хранение
Способность животных усваивать крахмал зависит от их способности вырабатывать амилазу. Было показано, что все виды рыб секретируют -амилазу. Также было продемонстрировано, что активность этого фермента была наибольшей у травоядных животных.У плотоядных, таких как радужная форель и морской окунь, амилаза в основном имеет панкреатическое происхождение, тогда как у травоядных этот фермент широко распространен по всему пищеварительному тракту. У Tilapia mossambica было показано, что поджелудочная железа является местом наибольшей активности амилазы, за которым следует верхний отдел кишечника. Хотя было показано, что переваривание крахмала и декстрина плотоядной радужной форелью постепенно снижается по мере того, как уровни углеводов превышают 20-процентный уровень, рыба может эффективно использовать до 60 процентов глюкозы, сахарозы или лактозы в рационе.Это демонстрирует, что вопреки более раннему мнению, плотоядные рыбы способны эффективно использовать простые углеводы в качестве основного источника энергии.
Кристаллическая структура крахмала, по-видимому, также влияет на его атаку амилазой, о чем свидетельствует двукратное увеличение содержания метаболизируемой энергии в полностью приготовленной (желатинизированной) кукурузе при испытаниях кормления канальным сомом. Также было показано, что радужная форель имеет более высокую толерантность к углеводам (присутствующим в виде пшеничного крахмала) в рационе при приготовлении.Процесс желатинизации включает в себя тепло и воду. Если водную суспензию крахмала нагреть, гранулы не изменят внешний вид, пока не будет достигнута определенная критическая температура. В этот момент некоторые гранулы крахмала набухают и одновременно теряют свою кристалличность. Критическая температура - это температура, при которой водородные связи молекулы крахмала ослабляются, чтобы обеспечить полную гидратацию, что приводит к явлению, известному как «набухание».
Альфа-амилаза способствует более или менее случайной фрагментации молекулы крахмала путем гидролиза по глюкозидным связям a -D- (l® 4) во внутренней и внешней цепях соединения.Результатом полного гидролиза амилозного компонента являются мальтоза и D-глюкоза, в то время как амилопектиновый компонент восстанавливается до мальтозы, D-глюкозы и разветвленных предельных декстринов. Вследствие этих паттернов действия -амилазы на крахмал, необходимы другие ферменты для полного гидролиза крахмала до D-глюкозы у рыб. В связи с этим было продемонстрировано, что даже плотоядный морской лещ обладает способностью переваривать мальтозу. С другой стороны, не было показано, что целлюлаза и -галактозидаза секретируются рыбами, хотя целлюлаза бактериального происхождения присутствует в кишечнике большинства видов карпов.Недостаток -галактозидазы может частично объяснить слабую реакцию рыбы на диетическую соевую муку, которая содержит значительные уровни галактозидных олигосахаридов рафинозы и стахиозы. Как указывалось ранее, эти олигосахариды действительно подвергаются ферментативному гидролизу в процессе прорастания с образованием галактозы и сахарозы. Следовательно, может показаться, что пищевая ценность соевого шрота будет увеличена, если сначала преобразовать большую часть этого неперевариваемого крахмала. Этого можно достичь, замачивая бобы на 48 часов перед переработкой для производства муки.Следует также отметить, что питательная ценность зернобобовых и других семян бобовых также может быть улучшена для рыбы, поскольку олигосахариды составляют большую часть углеводов в семенах бобовых.
Данных о всасывании глюкозы рыбами немного. Работа с золотыми рыбками показала, что активный транспорт глюкозы связан с транспортом Na + , как и у большинства млекопитающих. Обычно считается, что абсорбция происходит на поверхности слизистой оболочки кишечных клеток. Моносахариды, образующиеся в результате переваривания углеводов, состоят в основном из глюкозы, фруктозы, галактозы, маннозы, ксилозы и арабинозы.Хотя скорость поглощения этих сахаров была определена для многих наземных млекопитающих, аналогичная информация для рыб отсутствует.
Глюкоза, по-видимому, не является лучшим источником энергии для рыбы по сравнению с белком или жиром, хотя легкоусвояемые углеводы экономят белок для построения тканей. Кроме того, в отличие от млекопитающих, гликоген не является значительным хранилищем энергии, несмотря на свидетельства активного и обратимого пути Эмдена-Мейерхоффа у рыб. Более эффективный метаболизм аминокислот по сравнению с глюкозой для получения энергии может быть связан со способностью рыб выводить азотсодержащие отходы в виде аммиака из своих жабр без высоких затрат энергии на преобразование отходов в мочевину.
3.2 Другие факторы, влияющие на метаболизм
Помимо генетической адаптации, климатические факторы также играют важную роль в углеводном обмене у рыб. Акклиматизация рыб, по сути, отражает акклиматизацию ферментов, поскольку способность животного выживать во многом зависит от его способности выполнять нормальные метаболические функции. Некоторые ферменты метаболической акклиматизации хорошо компенсируются, а другие нет. Ферменты, связанные с высвобождением энергии (ферменты гликолиза, пентозного шунта, цикла трикарбоновых кислот, транспорта электронов и окисления жирных кислот), демонстрируют температурную компенсацию, тогда как те ферменты, которые в основном связаны с деградацией продуктов метаболизма, демонстрируют плохую или обратную компенсацию (см. Таблицу 1). .
Таблица 1 Ферменты, подлежащие метаболической акклиматизации 1/
| Ферменты с компенсацией | Ферменты с обратной компенсацией или без нее |
| фосфофруктокиназа | каталаза |
| альдолаза | пероксидаза |
| лактодегидрогеназа | кислая фосфатаза |
| 6-фосфоглюконатдегидрогеназа | оксидаза D-аминокислот |
| янтарная дегидрогеназа | Mg-ATP азе |
| яблочная дегидрогеназа | холинацетилтрансфераза |
| цитохромоксидаза | ацетилхолинэстераза |
| сукцинат-цитохром С редуктаза | щелочная фосфатаза |
| НАД-цитохром С редуктаза | аллантоиназа |
| аминоацилтрансфераза | уриказа |
| Na-K-АТФаза | амилаза |
| протеаза | липаза |
| яблочный фермент | |
| глюкозо-6-фосфатдегидрогеназа |
1/ Взято из: Сравнительная физиология животных, под редакцией К.Л. Проссер, 1973
Интересно отметить, что два ключевых фермента, участвующих в метаболизме углеводов, амилаза и глюкозо-6-фосфатдегидрогеназа, вместе с ферментом, участвующим в переваривании жиров, липазой, не имеют температурной компенсации. Неизвестно, связано ли это каким-либо образом с прекращением кормления рыб при низких температурах. Молекулярный механизм термической акклиматизации не совсем понятен и может состоять из изменений в синтезе или количествах данного фермента.Различия в кинетике, изменения в пропорции изоферментов, подходящих для определенных температур, и изменения кофакторов, таких как липиды, коферменты, или других факторов, таких как pH и ионы, могут иметь важное значение для адаптации животного к изменениям температуры.
3.3 Преобразование энергии
Несмотря на межвидовые различия в переносимости пищевых углеводов, обычно считается, что основной конечный продукт переваривания углеводов, глюкоза, метаболизируется способом, преобладающим во всех клетках, т.е.е., через , обратимый путь Эмдена-Мейерхоффа. На этом пути глюкоза имеет только одну главную судьбу: фосфорилирование до глюкозо-6-фосфата. Основные метаболические превращения изображены следующим образом:
Обратимые стрелки показывают стадию или стадии реакции, катализируемые теми же ферментами в в обоих направлениях.
Пунктирные стрелки показывают реакции на многих промежуточных этапах.
Парные сплошные стрелки показывают разные ферменты, участвующие в двух направлениях. реакции.
(по материалам: Principles of Biochemistry, A. White, et al. , al ., 1978)
Все превращения происходят с потерей свободной энергии. Таким образом, образование двух молей лактата из глюкозо-6-фосфата происходит с изменением свободной энергии D G o = -22000 кал / моль. Конечный результат - образование четырех молекул АТФ. Функциональное обращение этой трансформации может происходить только через другой последовательности, требующей ввода шести молекул АТФ на моль извлеченного глюкозо-6-фосфата.
Клетки не хранят глюкозу или глюкозо-6-фосфат. Легко доступная форма хранения - это гликоген, который производится из глюкозо-1-фосфата одним путем и возвращается другим. Хотя в клетках млекопитающих глюкозо-6-фосфат трансформируется в жирные кислоты, такое превращение, по-видимому, не происходит у рыб. Исследования с карпом показывают, что предшественником липогенеза является цитрат, образующийся, когда аминокислоты активно метаболизируются в цикле трикарбоновых кислот.
Основной формой полезной энергии во всех клетках является АТФ. В большинстве клеток эта валюта энергии генерируется окислением НАДН митохондриальными системами транспорта электронов. Восстановители NAD + для этого процесса представляют собой промежуточные продукты, полученные из цикла TCA и жирных кислот. Энергетический выход глюкозы в дыхательной системе можно суммировать в следующей последовательности реакций:
| Реакция | Выход ATP | |
| 1.глюкоза® фруктозо-1,6-дифосфат | -2 | |
| 2. 2-триозофосфат® 2,3-фосфоглицериновый кислота | +2 | |
| 3. 2 НАД + ® 2 НАДН® 2 NAD + | +6 | |
| 4.2 фосфоенолпируват® 2 пировиноградная кислота | +2 | |
| 5. 2 пировиноградная кислота® 2 ацетил-КоА + 2 CO 2 | ||
| 2 НАД + ® 2 НАДН® 2 НАД 2 | +6 | |
| 6.2 Ацетил CoA® 4 CO 2 | +24 | |
| Всего: | ||
| C 6 H 12 O 6 + 6O 2 ® 6 CO 2 + 6 H 2 O | +38 | |
Проссер, К.Л. (ред.), 1973 г. Сравнительная физиология животных. Филадельфия, W.B. Компания Saunders, 1011 стр. 3-е изд.
White, A., et al. , al. ., 1978 г., Принципы биохимии. Нью-Йорк, McGraw-Hill Book Company, 1492 стр. 6-е изд.
.Учебное пособие по
: Глава 8 Нуклеотиды и нуклеиновые кислоты - NUS
Вопросы с множественным выбором
- Некоторые основы Страницы: 271-273 Сложность: 1 Ответ: E Соединение, состоящее из рибозы, связанной N-гликозидной связью с N-9 аденина, составляет:
A) дезоксирибонуклеозид. Б) пуриновый нуклеотид. В) пиримидиновый нуклеотид. Г) аденозинмонофосфат. Е) аденозин.
- Некоторые основы Страница: 273 Сложность: 1 Ответ: E Основной компонент РНК, но не ДНК:
А) аденин.Б) цитозин. В) гуанин. Г) тимин. Д) урацил.
- Некоторые основы Страница: 273 Сложность: 1 Ответ: A Разница между рибонуклеотидом и дезоксирибонуклеотидом:
A) дезоксирибонуклеотид имеет -H вместо -OH у C-2. B) дезоксирибонуклеотид имеет конфигурацию; рибонуклеотид имеет -конфигурацию у C-1. C) рибонуклеотид имеет дополнительный -OH при C-4. D) рибонуклеотид обладает большей структурной гибкостью, чем дезоксирибонуклеотид. E) рибонуклеотид представляет собой пиранозу, дезоксирибонуклеотид представляет собой фуранозу.
- Некоторые основы Страница: 273 Сложность: 2 Ответ: D Что из следующего верно для пентоз, обнаруженных в нуклеиновых кислотах?
A) C-5 и C-1 пентозы присоединены к фосфатным группам. B) C-5 пентозы присоединен к азотистому основанию, а C-1 - к фосфатной группе. C) Связь, которая соединяет азотистые основания с пентозами, представляет собой O-гликозидную связь. D) Пентозы всегда находятся в формах-фуранозы. E) Прямоцепные и кольцевые формы подвергаются постоянному взаимопревращению.
- Некоторые основы Страницы: 274-275 Сложность: 2 Ответ: E Фосфодиэфирные связи, которые связывают соседние нуклеотиды как в РНК, так и в ДНК:
A) всегда связывайте A с T, а G с C. Б) подвержены щелочному гидролизу. C) не заряжены при нейтральном pH. Г) образуют между плоскими кольцами соседних оснований. E) присоединить 3'-гидроксил одного нуклеотида к 5'-гидроксилу следующего.
- Некоторые основы Страницы: 274-275 Сложность: 2 Ответ: A Фосфодиэфирная связь, соединяющая соседние нуклеотиды в ДНК:
A) ионно связывается с ионами металлов, полиаминами и белками.Б) заряжен положительно. C) подвержен щелочному гидролизу. D) Связывает C-2 одной базы с C-3 следующей. E) связывает C-3 дезоксирибозы с N-1 тимина или цитозина.
- Некоторые основы Страница: 275 Сложность: 2 Ответ: D Щелочной гидролиз РНК не дает:
А) 2 '- AMP. Б) 2 ', 3'-цГМФ. В) 2'-CMP. Г) 3 ', 5'-цАМФ. E) 3'-UMP.
- Некоторые основы Страница: 276 Сложность: 2 Ответ: B ДНК-олигонуклеотид, сокращенно pATCGAC: .
A) имеет 7 фосфатных групп.B) имеет гидроксил на своем 3'-конце. C) имеет фосфат на его 3'-конце. D) имеет букву A на 3 'конце. E) нарушает правила Чаргаффа.
- Некоторые основы Страница: 276 Сложность: 2 Ответ: D Для олигорибонуклеотида pACGUAC:
A) нуклеотид на 3'-конце имеет фосфат на 3'-гидроксиле. Б) нуклеотид на 3'-конце представляет собой пурин. C) нуклеотид на 5'-конце имеет 5'-гидроксил. D) нуклеотид на 5'-конце имеет фосфат на 5'-гидроксиле. E) нуклеотид на 5'-конце представляет собой пиримидин.
- Структура нуклеиновой кислоты Страница: 278 Сложность: 1 Ответ: B Эксперимент Эйвери, Маклауда и Маккарти, в котором невирулентные бактерии стали вирулентными по преобразованию было значительным, поскольку показало, что:
А) бактерии могут подвергаться трансформации. Б) гены состоят только из ДНК. В) мыши более восприимчивы к пневмонии, чем люди. Г) пневмонию можно вылечить трансформацией. Д) вирулентность определяется генетически.
- Структура нуклеиновой кислоты Страница: 278 Сложность: 1 Ответ: E Правила Чаргаффа гласят, что в типичной ДНК:
A) A = G.Б) А = С. В) А = U. Г) А + Т = G + C. E) A + G = T + C.
- Структура нуклеиновой кислоты Страница: 278 Сложность: 1 Ответ: A Исходя из правил Чаргаффа, какие из следующего являются возможными базовыми композициями для двухцепочечных ДНК?
% A% G% C% T% U А) 5 45 45 5 0 Б) 20 20 20 20 20 В) 35 15 35 15 0 Г) все вышеперечисленное E) ничего из вышеперечисленного
- Структура нуклеиновой кислоты Страницы: 278-280 Сложность: 2 Ответ: A В структуре ДНК Уотсона-Крика:
A) отсутствие 2'-гидроксильных групп позволяет основаниям располагаться перпендикулярно оси спирали.Б) содержание аденина в одной цепи должно равняться содержанию тимина в той же цепи. В) нуклеотиды расположены в А-форме. D) содержание пуринов (доля оснований, которые являются пуринами) должно быть одинаковым в обеих цепях. Д) две пряди параллельны.
- Структура нуклеиновой кислоты Страницы: 278-280 Сложность: 2 Ответ: E В модели структуры ДНК Уотсона-Крика:
A) обе нити проходят в одном направлении, 3 ' 5'; они параллельны. Б) фосфатные группы выступают к середине спирали, где они защищены от взаимодействие с водой.C) T может образовывать три водородные связи либо с G, либо с C в противоположной цепи. D) расстояние между сахарной основой двух нитей достаточно велико, чтобы вместить либо два пурина, либо два пиримидина. E) расстояние между двумя соседними основаниями в одной нити составляет около 3,4 Å.
- Структура нуклеиновой кислоты Страницы: 278-280 Сложность: 2 Ответ: C Что из следующего не относится ко всей природной ДНК?
A) Дезоксирибозные звенья связаны 3 ', 5'-фосфодиэфирными связями.Б) Количество A всегда равно количеству T. C) Отношение A + T / G + C постоянно для всех природных ДНК. D) Две дополнительные цепи антипараллельны. E) Две водородные связи образуются между A и T.
- Структура нуклеиновой кислоты Страницы: 278-280 Сложность: 2 Ответ: E В модели структуры ДНК Уотсона-Крика (теперь называемой ДНК B-формы):
A) пурин в одной цепи всегда водородными связями с пурином в другой цепи. B) Пары A – T имеют три водородные связи. C) Пары G – C имеют две водородные связи.D) 5 'концы обеих нитей находятся на одном конце спирали. E) основания занимают внутреннюю часть спирали.
- Структура нуклеиновой кислоты Страницы: 278-280 Сложность: 2 Ответ: D Двойная спираль ДНК в B-форме стабилизирована:
A) ковалентные связи между концом 3 'одной нити и концом 5' другой. Б) водородная связь между фосфатными группами двух расположенных рядом нитей. В) водородные связи между рибозами каждой нити. D) неспецифическое взаимодействие при укладке оснований между двумя соседними основаниями в одной цепи.E) взаимодействия рибозы с плоскими парами оснований.
- Структура нуклеиновой кислоты Страница: 280 Сложность: 2 Ответ: C В нуклеотидах и нуклеиновых кислотах син- и антиконформации относятся к:
А) основные стереоизомеры. Б) вращение вокруг фосфодиэфирной связи. В) вращение вокруг сахарной основы. Г) сахарная морщинка. Д) стереоизомеры сахаров.
- Структура нуклеиновой кислоты Страница: 284-286 Сложность: 1 Ответ: D Какие из следующих возможных оснований для одноцепочечной РНК?
% A% G% C% T% U А) 5 45 45 0 5 Б) 25 25 25 0 25 В) 35 10 30 0 25 Г) все вышеперечисленное E) ничего из вышеперечисленного
- Структура нуклеиновой кислоты Страницы: 284-286 Сложность: 2 Ответ: C Двухцепочечные области РНК:
A) менее стабильны, чем двухцепочечные участки ДНК.B) можно наблюдать в лаборатории, но, вероятно, не имеет биологического значения. C) могут образовываться между двумя самокомплементарными участками одной и той же цепи РНК. Г) не встречаются. E) две нити расположены параллельно (в отличие от ДНК, которые антипараллельны).
- Химия нуклеиновых кислот Страницы: 287-288 Сложность: 2 Ответ: B Когда двухцепочечная ДНК нагревается до нейтрального pH, какое изменение не происходит?
A) Поглощение ультрафиолетового (260 нм) света увеличивается.Б) Ковалентная N-гликозидная связь между основанием и пентозой разрывается. В) Винтовая структура раскручивается. D) Водородные связи между A и T разрываются. E) Вязкость раствора уменьшается.
- Химия нуклеиновых кислот Страницы: 288-289 Сложность: 2 Ответ: B Какой из следующих дезоксиолигонуклеотидов будет гибридизоваться с ДНК, содержащей последовательность (5 ') AGACTGGTC (3')?
А) (5 футов) CTCATTGAG (3 фута) B) (5 ') GACCAGTCT (3') C) (5 ') GAGTCAACT (3') D) (5 ') TCTGACCAG (3') E) (5 футов) TCTGGATCT (3 дюйма)
- Химия нуклеиновых кислот Страницы: 288-289 Сложность: 2 Ответ: D Рибонуклеотидный полимер (5 ') GTGATCAAGC (3') может образовывать только двухцепочечную структуру с:
A) (5 футов) CACTAGTTCG (3 фута).B) (5 ') КАКУАГУУК (3'). C) (5 ') CACUTTCGCCC (3'). D) (5 ') GCTTGATCAC (3'). E) (5 ') GCCTAGTTUG (3').
- Химия нуклеиновых кислот Страница: 288 Сложность: 2 Ответ: E По сравнению с двойными спиралями ДНК-ДНК, стабильность спиралей ДНК-РНК и РНК-РНК составляет:
А) ДНК-ДНК> ДНК-РНК> РНК-РНК. Б) ДНК-ДНК> РНК-РНК> ДНК-РНК. В) РНК-ДНК> РНК-РНК> ДНК-ДНК. Г) РНК-РНК> ДНК-ДНК> ДНК-РНК. E) РНК-РНК> ДНК-РНК> ДНК-ДНК.
- Химия нуклеиновых кислот Страница: 290 Сложность: 1 Ответ: E В лабораторных условиях известно, что несколько факторов вызывают изменение химической структуры ДНК.В фактор (ы), который может быть важным в живой клетке:
А) тепло. Б) низкий pH. В) кислород. Г) УФ-свет. E) как C, так и D.
- Химия нуклеиновых кислот Страница: 291 Сложность: 2 Ответ: B Соединения, образующие азотистую кислоту (например, нитриты, нитраты и нитрозамины), изменяют ДНК. молекул по:
А) разрыв фосфодиэфирных связей. Б) дезаминирование оснований. В) депуринизация. Г) образование димеров тимина. E) преобразование A T.
- Химия нуклеиновых кислот Страницы: 292-294 Сложность: 2 Ответ: D При секвенировании ДНК методом Сэнгера (дидезокси):
A) радиоактивный дидезокси-АТФ включается в каждую из четырех реакционных смесей перед ферментативной синтез комплементарных цепей.Б) специальные ферменты используются для разрезания синтезированной ДНК на мелкие кусочки, которые затем разделены электрофорезом. C) дидезоксинуклеотиды должны присутствовать в больших количествах для получения длинных участков последовательности ДНК. D) роль дидезокси CTP заключается в том, чтобы иногда останавливать ферментативный синтез ДНК, где Gs возникают в прядях шаблона. E) цепь матричной ДНК радиоактивна.
- Некоторые основы Страницы: 271-277 Сложность: 1 Сопоставьте тип облигации с ролью ниже:
Bond_type Роль (а) фосфодиэфир _ связывает основание с пентозой в нуклеотиде (b) N-гликозид _ соединяет соседние нуклеотиды в одну цепь (c) сложный фосфатный эфир _ соединяет комплементарные нуклеотиды двумя пряди (d) водород _ разница между нуклеозидом и нуклеотид Ответ: б; а; d; в
- Структура нуклеиновой кислоты Страницы: 271-277 Сложность: 2 Соединения, содержащие азотистое основание, сахар и фосфатную группу, называются (а) _____ .Два пурина, обнаруженные в ДНК: (b) __ и __ . Пиримидин присутствует во всей ДНК, но только в некоторых РНК. (в) _____ . В ДНК пара оснований (d) - удерживается вместе тремя водородными связями; пара оснований (e) - имеет только две такие связи.
Ответ: (а) нуклеотиды; (б) аденин и гуанин; (c) тимидин; (d) G – C; (e) A – T
- Структура нуклеиновой кислоты Страница: 277 Сложность: 2 Изобразите структуру пары оснований аденин-тимин или гуанин-цитозин, как показано на Двойная спиральная структура ДНК Уотсона-Крика.Включите все водородные связи.
Ответ: (См. Рис. 8-11. Стр. 277.)
- Структура нуклеиновой кислоты Страница: 277 Сложность: 2 Изобразите структуры аденина и тимина с водородными связями.
Ответ: (См. Рис. 8-11, стр. 277.)
- Структура нуклеиновой кислоты Страница: 278 Сложность: 3 Кратко опишите экспериментальные доказательства Эйвери, Маклауда и Маккарти, что ДНК является генетическим материал.
Ответ: Эйвери и др. показали, что ДНК, выделенная из вирулентной (болезнетворной) бактерии (Streptococcus pneumoniae) при смешивании с живыми клетками невирулентного штамма этой бактерии, предоставили генетические инструкции по трансформации невирулентного штамма в вирулентный штамм.
- Структура нуклеиновой кислоты Страницы: 278-279 Сложность: 3 Состав (мольная доля) одной из нитей двойной спирали ДНК составляет [A] = 0,3 и [G] = 0,24. Если возможно, рассчитайте следующее. Если невозможно, напишите «Я».
Для той же пряди: [T] = (a) __ [C] = (b) __ [T] + [C] = (c) __ Для другой пряди: [A] = (d) __ [T] = (e) __ [A] + [T] = (f) __ [G] = (g) __ [C] = (h) __ [G] + [C] = (i) ____
Ответ: (а) Я; (б) я; (в) 0.46; (г) я; е) 0,3; (f) я; (g) я; (з) 0,24; (i) Я
- Структура нуклеиновой кислоты Страница: 279 Сложность: 2 Какова приблизительная длина молекулы ДНК (в форме B), содержащей 10 000 пар оснований?
Ответ: 3,4 Å 10,000 = 34,000 Å = 3,4 мкм. (См. Стр. 338.)
- Структура нуклеиновой кислоты Страница: 279 Сложность: 2 Кратко опишите, что имеется в виду, говоря, что две цепи ДНК комплементарны.
Ответ: нуклеотидные последовательности комплементарных цепей таковы, что всякий раз, когда A встречается в одной цепи, в другой цепи есть T, с которой она может образовывать пару оснований с водородными связями.Где бы C встречается в одной нити, а G - в другой. A - база, дополнительная к T, а C - база дополняет G.
-
Структура нуклеиновой кислоты Страницы: 280-281 Сложность: 2 Одним предложением определите наиболее очевидное структурное различие между А-образной формой (Ватсон-Крик) ДНК и Z-форма ДНК. Ответ: А-форма ДНК представляет собой правую спираль; Z-форма ДНК представляет собой левую спираль. (См. Рис. 10-19, стр. 338.)
-
Структура нуклеиновой кислоты Страницы: 281-282 Сложность: 2 Напишите двухцепочечную последовательность ДНК, содержащую палиндром из шести нуклеотидов.
Ответ: Любая двухцепочечная последовательность, имеющая форму: 1-2-3-4-5-6-6'-5'-4'-3'-2'-1 ' 1 '-2'-3'-4'-5'-6'-6-5-4-3-2- где каждое число и его простое число представляют правильно спаренные основания (A с T, C с G), все вдоль двухцепочечной молекулы.
- Химия нуклеиновых кислот Страницы: 290-291 Сложность: 2 Каково основное влияние ультрафиолетового излучения на ДНК?
Ответ: УФ-излучение вызывает образование димера между соседними основаниями тимина на одном и том же Нить ДНК.Это приводит к изгибу двойной спирали в этом месте.
- Другие функции нуклеотидов Страницы: 296-298 Сложность: 2 Объясните, как нуклеозидтрифосфаты (например, АТФ) действуют как переносчики химической энергии.
Ответ: В этих молекулах существует фосфорно-ангидридная связь между фосфатами. Гидролиз этой связи (т. е. АТФ с АДФ и фосфатом) представляет собой сильно экзэргоническую реакцию, которая приводит к высвобождение большого количества свободной энергии. И наоборот, вклад большого количества свободной энергии равен требуется для синтеза связи; я.е. для фосфорилирования АДФ с образованием АТФ. Таким образом, Синтез АТФ приводит к накоплению энергии, которая может высвобождаться при гидролизе.
.Структура нуклеиновой кислоты- вопросы и ответы по биохимии
перейти к содержанию Меню- Дом
- разветвленных MCQ
- Программирование
- CS - IT - IS
- CS
- IT
- IS
- ECE - EEE - EE
- ECE
- EEE
- EE
- Гражданский
- Механический
- Химическая промышленность
- Металлургия
- Горное дело
- Приборы
- Аэрокосмическая промышленность
- Авиационная
- Биотехнологии
- Сельское хозяйство
- Морской
- MCA
- BCA
- Тест и звание
- Sanfoundry Tests
- Сертификационные испытания
- Тесты для стажировки
- Занявшие первые позиции
- Конкурсы
- Стажировка
- Обучение