Какие продукты не любят лямблии
Диета при лямблиозе: рецепты, меню при лечении лямблий у детей и взрослых
Закрыть- Болезни
- Инфекционные и паразитарные болезни
- Новообразования
- Болезни крови и кроветворных органов
- Болезни эндокринной системы
- Психические расстройства
- Болезни нервной системы
- Болезни глаза
- Болезни уха
- Болезни системы кровообращения
- Болезни органов дыхания
- Болезни органов пищеварения
- Болезни кожи
- Болезни костно-мышечной системы
- Болезни мочеполовой системы
- Беременность и роды
- Болезни плода и новорожденного
- Врожденные аномалии (пороки развития)
- Травмы и отравления
- Симптомы
- Системы кровообращения и дыхания
- Система пищеварения и брюшная полость
- Кожа и подкожная клетчатка
- Нервная и костно-мышечная системы
- Мочевая система
- Восприятие и поведение
- Речь и голос
- Общие симптомы и признаки
- Отклонения от нормы
- Диеты
- Снижение веса
- Лечебные
- Быстрые
- Для красоты и здоровья
- Разгрузочные дни
- От профессионалов
- Монодиеты
- Звездные
- На кашах
- Овощные
- Детокс-диеты
- Фруктовые
- Модные
- Для мужчин
- Набор веса
- Вегетарианство
- Национальные
- Лекарства
- Антибиотики
- Антисептики
- Биологически активные добавки
- Витамины
- Гинекологические
- Гормональные
- Дерматологические
- Диабетические
- Для глаз
- Для крови
- Для нервной системы
- Для печени
- Для повышения потенции
- Для полости рта
- Для похудения
- Для суставов
- Для ушей
- Желудочно-кишечные
- Кардиологические
- Контрацептивы
- Мочегонные
- Обезболивающие
- От аллергии
- От кашля
- От насморка
- Повышение иммунитета
- Противовирусные
- Противогрибковые
- Противомикробные
- Противоопухолевые
- Противопаразитарные
- Противопростудные
- Сердечно-сосудистые
- Урологические
- Другие лекарства
- Врачи
- Клиники
- Справочник
- Аллергология
- Анализы и диагностика
- Беременность
- Витамины
- Вредные привычки
- Геронтология (Старение)
- Дерматология
- Дети
- Женское здоровье
- Инфекция
- Контрацепция
- Косметология
- Народная медицина
- Обзоры заболеваний
- Обзоры лекарств
- Ортопедия и травматология
- Питание
- Пластическая хирургия
- Процедуры и операции
- Психология
- Роды и послеродовый период
- Сексология
- Стоматология
- Травы и продукты
- Трихология
- Другие статьи
- Словарь терминов
- [А] Абазия .. Ацидоз
- [Б] Базофилы .. Богатая тромбоцитами плазма
- [В] Вазопрессин .. Выкидыш
- [Г] Галлюциногены .. Грязи лечебные
- [Д] Деацетилазы гистонов .. Дофамин
- [Ж] Железы .. Жиры
- [И] Иммунитет .. Искусственная кома
- [К] Каверна .. Кумарин
- [Л] Лапароскоп .. Лучевая терапия
- [М]
Giardia lamblia: морфология, жизненный цикл, патогенез, клинические проявления, лабораторная диагностика и лечение

- Giardia lamblia также известна как интестинилис или G.duodenalis.
- Впервые его заметил Антоний фон Левенгук (1681 г.), исследуя свой стул, а Ламби (1859 г.) описал паразита и назвал его Giardia labmlia
- Лямблии являются единственным кишечным жгутиком, вызывающим эндемическую и эпидемическую диарею у человека.
Место обитания:
- Населяет тонкую кишку человека
Морфология:
G. lamblia существует в двух морфологических формах - трофозоит и циста
-
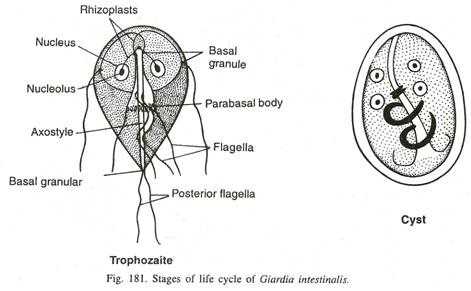
Трофозоит:
- Это стадия активного питания паразита, ответственная за колонизацию в кишечнике.
- Трофозоит имеет форму груши или теннисной ракетки с широким круглым передним концом и сужающимся задним концом.
- Его длина составляет 9–21 мкм, а ширина - 5–5 мкм.
- Дорсальная поверхность выпуклая, в то время как брюшная поверхность вогнутая, с присасывающим диском (липким диском), который действует как орган для прикрепления.
- За клейким диском находится пара больших изогнутых и поперечных срединных тел, уникальных для
- Он двусторонне симметричен, все органы тела парные. У них два срединных тела, два аксостиля, два ядра и четыре пары жгутиков.
- Каждое ядро состоит из большой центральной кариосомы, придающей паразиту характерный вид лица в окрашенном препарате.
- Цитоплазма однородная и мелкозернистая.
- Подвижность: типичная подвижность «опадающего листа».
-
Киста:
- Это инфекционная стадия паразита.
- Полностью созревшая киста имеет овальную или эллипсоидальную форму, длину 8–12 мкм и ширину 7–10 мкм.
- Киста окружена толстой стенкой кисты. Цитоплазма гранулирована и отделена от стенки кисты прозрачным пространством.
- Аксостиль лежит более или менее по диагонали.
- Киста содержит 4 ядра.
- Внутри цитоплазмы видны остатки жгутиков и края сосательного диска.
Жизненный цикл Giardia lamblia :
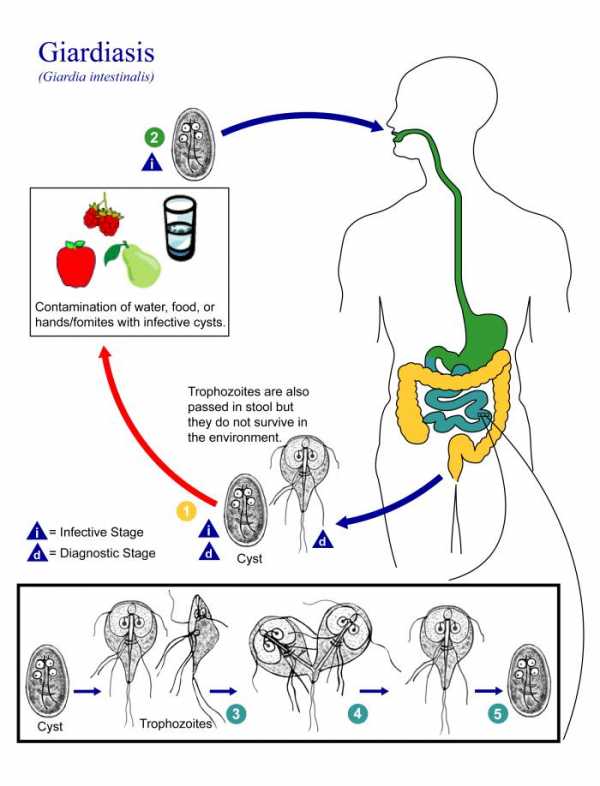
- Жизненный цикл G. lamblia прост и завершается у одного хозяина - человека. Промежуточный хост не требуется.
- Инфекция передается орально при попадании кисты через зараженную руку, воду или пищу.
- Эксцистация происходит в желудке и двенадцатиперстной кишке в присутствии кислоты желудочного сока, ферментов поджелудочной железы (химотрипсина и трипсина).Для эксцистанции требуется кислая среда с pH 1,3-2,7.
- Каждая киста эксцистируется с образованием двух трофозоитов в двенадцатиперстной кишке в течение 30 минут после приема внутрь.
- Эти трофозоиты размножаются в кишечнике за счет бунарного деления. Затем они прикрепляются к энтероцитам через вентральную присоску, возможно, через поверхностный связывающий маннозу лектин, присутствующий на поверхности трофозоитов.
- Некоторые трофозоиты затем переходят в толстую кишку, где они снова энцифицируются в присутствии нейтрального pH и солей желчных кислот.
- Процесс энцистации начинается с появления секреторных пузырьков, специфичных для энцистации (ESV) в цитоплазме трофозоитов, с последующим образованием стенки кисты в течение 15 часов.
- В течение 24 часов после появления ESV трофозоит покрывается этими белками стенки кисты, что приводит к образованию кисты.
- Формирование кисты начинается с укорочения жгутиков, за которым следует конденсация цитоплазмы и, наконец, секреция толстой стенки кисты хилина.
- Затем эти инцистированные трофозоиты претерпевают другую фазу деления ядра и образуют четырехъядерные зрелые кисты.
- Цисты, являющиеся инфекционной формой паразита, выделяются с фекалиями, и жизненный цикл повторяется.
Патогенез и патология:
Способ передачи:
- Человек - главный резервуар лямблий.
- Заражение приобретено:
- Проглатывание зараженных продуктов питания и воды
- Передача от человека к человеку в результате плохой гигиены в детских садах, домах престарелых, психиатрических лечебницах
- Передача половым путем - орально-анальный и орально-генитальный секс
- Лица с ослабленным иммунитетом, такие как пациенты со СПИДом, Х-сцепленная гаммаглобулинемия, пациенты с белково-энергетической недостаточностью, более восприимчивы к лямблиозу
Факторы вирулентности:
- Цитоскелет:
- Лямблии содержат цитоскелецию микротрубочек (МТ), которая необходима для подвижности, прикрепления, внутриклеточного транспорта, деления клеток и энцистации / эксцистации.
- Кисты:
- Цисты устойчивы и ответственны за передачу паразитов
Патогенез Giardia lamblia :
- Лямблии - кишечные паразиты, неинвазивные.
- Как только происходит эксцистация, трофозоиты высвобождаются, и они используют свои жгутики, чтобы «плавать» к покрытой микроворсинками поверхности двенадцатиперстной и тощей кишки, где они прикрепляются к энтероцитам с помощью своего адгезивного диска.
- Лектины, присутствующие на поверхности Giardia , связываются с рецептором, присутствующим на поверхности энтероцитов. Этот процесс прикрепления повреждает микроворсинки, которые мешают всасыванию пищи ворсинками.
- Быстрое размножение трофозоитов в конечном итоге создает физические барьеры между энтероцитами и просветом кишечника, что еще больше препятствует усвоению пищи. Этот процесс приводит к повреждению энтероцитов, атропии ворсинок, гиперплазии крипт, повышенной проницаемости кишечника и повреждению щеточной границы, что вызывает снижение секреции дисахаридных ферментов.
- Лектины и другие цитопатические вещества, выделяемые паразитами, также вызывают косвенное повреждение кишечного эпителия.
- Трофозоиты не проникают в окружающие ткани и не проникают в кровоток. Таким образом, инфекция обычно ограничивается просветом кишечника.
- Лямблиоз приводит к снижению абсорбции воды и глюкозы в тонких кишках, а повреждение кишечного эпителия приводит к нарушению всасывания электролитов и жидкостей, что приводит к осмотической диарее, известной как лямблиоз.
Клиническое проявление Giardia lamblia :
- Инкубация от 1 до 3 недель
- В большинстве случаев инфекция протекает бессимптомно.
- Симптоматическая инфекция чаще встречается у детей, чем у взрослых, из-за их более низкого иммунитета.
-
Острый лямблиоз:
- Характеризуется острой водянистой диареей, спазмами в животе, вздутием живота и метеоризмом. Иногда тошнота, рвота, лихорадка, сыпь или запор у некоторых.
- Гной, кровь и слизь в стуле не обнаруживаются.
- Состояние сохраняется 5-7 дней.
-
Хронический лямблиоз:
- Симптомы включают хроническую диарею с мальабсорбцией жира (стеаторея) и мальабсорбцией витамина А, белка и D-ксилозы, потерю веса, недомогание, тошноту, анорексию
- Выпуклость живота, тонкие конечности и задержка роста - наиболее частые признаки у детей.
- Длится несколько недель
- Внекишечные области встречаются редко, иногда крапивница и реактивный артрит наблюдаются в редких случаях
-
Осложнение:
- У взрослых, синдром мальабсорбции и потеря веса
- У детей задержка роста, отсроченные вехи достижения
- Лямблиоз - это заболевание, которое проходит самостоятельно, и прогрессирование до хронического состояния наблюдается только у 5% инфицированных людей, и смерть встречается редко.
Лабораторная диагностика Giardia lamblia :
-
Образец:
- Стул, дуоденальное содержимое, слизь, окрашенная желчью, биопсия двенадцатиперстной / тощей кишки
-
Осмотр стула:
Giardia lamblia - microbewiki
Сканирующая электронная микрофотография (SEM) трофозоита Giardia lamblia, показывающая внешние ультраструктурные детали.Страница микробной биореалмы рода Giardia lamblia
Классификация
Таксоны высшего порядка
| Домен | Эукариота |
|---|---|
| (ранг не указан) | Diplomonadida group |
| (ранг не указан) | Diplomonadida |
| Семья | Hexamitidae |
| Подсемейство | Giardiinae |
| Род | Лямблии |
Виды
Другие названия: Лямблии кишечные, Лямблии дуоденальные
Описание и значение
Giardia lamblia - жгутиковый микроаэрофильный микроорганизм, впервые обнаруженный Ван Левенгук в 1681 году, который обнаружил его в своем собственном диарейном стуле.Трофозоит G. lamblia , вегетативная, подвижная форма G. lamblia имеет грушевидную форму и уникальную морфологию, такую как два идентичных ядра, вентральный диск для прикрепления к кишечнику хозяина и жгутики [см. Также # Структура трофозоита ]. Киста является репродуктивной формой и состоит из защитной стенки кисты и четырех ядер.
Род Giardia выделен более чем из 40 видов. Известно, что вид G. lamblia инфицирует человека, млекопитающих, рептилий, птиц, коров, овец и свиней, в зависимости от штамма (Adam 2001).
G. lamblia - одна из основных причин заболеваний, передающихся через воду во всем мире (CDC, 2004), а инфекция приводит к лямблиозу (характеризующемуся мальабсорбцией и тяжелой диареей). Кишечная инфекция, вызванная лямблиозом, особенно серьезна в развивающихся странах, где возникновение лямблиоза во многом связано с загрязнением источников воды. В США G. lamblia было обнаружено как в питьевой, так и в рекреационной воде. В связи с высокой распространенностью лямблиоза г.lamblia представляет значительный интерес в сообществе клинических исследований. Однако патогенетические механизмы до конца не изучены. [См. Также #Pathology].
G. lamblia также имеет важное значение в эволюционной биологии. Считается, что из-за отсутствия митохондрий G. lamblia отличается от одной из самых ранних ветвей эукариот до начала эндосимбиотических отношений митохондрий (было предложено название царства «Archezoa»). Однако в настоящее время эта теория обсуждается.Новые доказательства в генетике, например, путем сравнения гена, кодирующего валил-тРНК-синтетазу (Hashimoto et al, 1998), и открытия сложных клеточных механизмов (таких как локализованный митохондриально-подобный механизм транспорта электронов (Lloyd, 2002). ) предполагают, что это могут быть более продвинутые организмы, которые когда-то имели митохондрии, но с тех пор потеряли их. Секвенирование полного генома G. lambdia в настоящее время находится в процессе, но полученные на данный момент результаты уже предоставили много информации о его эволюционной истории.
Структура генома
G. lamblia имеет два ядра одинакового размера, каждое из которых содержит полный геном одинакового размера (Yu et al, 2002). Оба ядра транскрипционно активны и реплицируются одновременно во время деления клетки. (Адам, 2001).
Геном G. lamblia состоит из 1,2 миллиона пар оснований, распределенных среди пяти линейных хромосом, каждая из которых фланкирована последовательностью теломер (5’TAGGG3 ’), сравнимой с другими эукариотами (Le Blancq et al, 1991).Упаковка хромосом осуществляется коровыми гистонами, но механизм немного отличается от других эукайот (Yee et al, 2007). [См. Также # Текущее исследование].
В геноме среднее содержание GC составляет 46% (проект генома Giardia lamblia). Полное картирование генома G. lamblia (штамм WB) в процессе (завершено на 95%, по состоянию на август 2007 г.) с использованием секвенирования «дробовиком». На данный момент предсказано 6488 открытых рамок считывания (ORF), из которых 4746 транскрибируются.
Тем не менее, исследования проводились на основе частичного генома.Его сравнение с другими видами эукариот открыло новую перспективу в эволюции этого вида.
Исследования по сравнению вышестоящей последовательности с генами, кодирующими цитоскелетный белок, обнаружили несколько согласованных последовательностей, например, богатую АТ область 5’AATTAAAAA3 ’, находящуюся между -30 и -70 выше. Эти последовательности были предложены в качестве промоторных областей для G. lamblia , хотя они относительно короче, чем у других эукариот. Показанная выше область, богатая АТ, важна для транскрипции.Кроме того, недавние исследования показали, что промоторов G. lamblia , в частности, AT-богатые, «TATA» -подобные области, вызывают продукцию стерильных антисмысловых транскриптов (Teodorvic et al, 2007).
Гены, кодирующие гомологи митохондриальных белков, таких как белок теплового шока 70 и шаперонин 60 (cpn60), были идентифицированы в геноме G. lamblia (Roger et al, 1998), и их экспрессия была обнаружена на протяжении всего жизненного цикла. за исключением времени эксцистанции.Это говорит о том, что Gardia не была премитохондриальной, но включала митохондриального предка, но с тех пор потеряла их. В том же контексте гены, кодирующие вали-тРНК синтетазу (ValRS), которые, как полагают, происходят из митохондрий, также были обнаружены в геноме G. lamblia (Hashimoto et al, 1998).
Структура ячеек
Структура трофозоита
Вегетативная форма G. lamblia , трофозоиты, имеют длину от 12 до 15 мкм и ширину от 5 до 9 мкм.Они обладают уникальными характеристиками морфологии: биядерная структура (каждая содержит полный набор генома (Yu, et al, 2002), четыре пары жгутиков, вентральный диск и срединное тело. Два ядра почти идентичны, не имеют видимые ядрышки и расположены симметрично.
Считается, что цитоскелет трофозоитов, в частности вентральный диск, играет главную роль в его прикреплении к кишечнику хозяина (Adam, 2001). Белки, которые находятся исключительно на вентральном диске, такие как актинин, альфа-актинин, миозин и тропомизин (Feely et al, 1982), а также лектин (Farthing et al, 1986), были предложены в качестве биохимических агентов, участвующих в привязанность.Четыре пары жгутиков расположены на передней правой, задней, каудальной и вентральной сторонах организма. Каждая состоит из 11 микротрубочек (включая две основные микротрубочки).
Центрин, белок цитоскелета, был обнаружен в трофозоитах. Исследования иммунофлоресценции выявили его присутствие во всех системах микротрубочек у трофозоитов, включая жгутики, срединные тела, базальные тела, а также липкий диск (Corrêa et al, 2004). Это предполагает, что центрин лямблий может быть белком, связанным с микротрубочками, и он может участвовать в подвижности и адгезии.
Специализированные мембранные структуры для транспорта электронов и генерации мембранного потенциала, как это видно в митохондриях (Lloyd 2002), также были обнаружены на цитоплазматической мембране трофозоитов, что указывает на их эволюционную связь с митохондриями.
Эндоплазматический ретикулум (ER) или структуры, подобные Гольджи, необходимые для транспортировки белков у эукариот, не наблюдались в трофозоитах G. lamblia с помощью микроскопа. Однако обычные ER-связанные белки, такие как chaperonin BiP, были идентифицированы в обширной мембраносвязанной системе (Soltys et al, 1996).Это предполагает наличие ER-подобной структуры с функциональностью, аналогичной таковой у других эукариот. Присутствие белка ARF, необходимого для отрастания пузырьков, также указывает на присутствие Golgi-подобной структуры (Lujan et al, 1995), хотя необходимы дальнейшие исследования.
Структура кисты
Микрофотография кисты лямблии лямблии, полученной методом окрашивания йодом.Киста G. lamblia имеет диаметр от 7 до 10 мкм и содержит четыре ядра. Киста покрыта 0.Стенка кисты толщиной 3 ~ 0,5 мкм. Стенка кисты состоит из двух слоев: внешнего нитевидного слоя и внутреннего перепончатого слоя (в дальнейшем состоящего из двух мембран). (Adam, 2001) После эксцистации одна киста превращается в двух трофозоитов.
Метаболизм
Цистовая форма имеет более низкую скорость метаболизма по сравнению с трофозоитами (примерно 10 ~ 20% трофозоитов), и ее стимулирует этанол (Paget et al, 1989).
Трофозоит G. lamblia является микроаэрофильным, в нем отсутствуют митохондрии и он зависит от цитохром-опосредованного окислительного фосфорилирования.(Адам, 2001). Они могут выполнять аэробный и анаэробный метаболизм в зависимости от концентрации кислорода в окружающей среде, хотя в основном они полагаются на ферментацию. К тому же брожение проводится даже в присутствии кислорода.
Аэробный и анаэробный метаболизм
Метаболическая активность трофозоитов зависит от концентрации кислорода и глюкозы. Когда кислород отсутствует, преобладает метаболизм глюкозы, а основным продуктом является аланин. В присутствии кислорода производство аланина подавляется (Papanastasiou et al, 1997).Когда кислород достигает концентрации 46 мкМ, CO2 и ацетат становятся основными продуктами метаболизма.
Общие ферменты для кислородной детоксикации, такие как супероксиддисмутаза, каталаза, пероксидаза, не были обнаружены в трофозоите G. lamblia (Brown, 1995). В G. lamblia управление окислительным стрессом, как полагают, осуществляется с помощью класса тиоредоксинредуктазы дисульфидредуктазы, который использует цистин в качестве первичного акцептора электронов (Brown et al., 1996b).Недавние открытия показали, что цитоплазматический фермент NAD (P) H менадионоксидоредуктаза (DT-диафораза) также играет роль в управлении окислительным стрессом (Sanchez et al, 2001) и может значительно усиливать рост при избыточной экспрессии (Lei et al, 2006). .
Углеводный обмен
Глюкоза необходима для роста трофозоитов, но не обязательна. Он вносит свой вклад в основной источник энергии, получаемый из углеводов. Путь метаболизма глюкозы у G. lamblia сходен с метаболизмом глюкозы у других эукариот, за исключением незначительных аспектов.Например, превращение фруктозо-6-фосфата в фруктозо-1,6-бисфосфат необратимо у большинства прокариот и эукариот, катализируется фосфофруктокиназой, требует АТФ и регулируется. Однако в G. lamblia этот процесс не регламентирован. И фосфофруктокиназа не является АТФ-зависимой, а напротив, пирофосфат-зависимой (Adam 2001, Michels et al, 2006).
Существуют доказательства того, что трофозоит использует гликоген в качестве запаса энергии для хранения энергии (Ladeira et al, 2005).
Аминокислотный обмен
G. lamblia способен синтезировать de novo только аланин (для энергетического метаболизма) и валин. Он полагается на извлечение всех других аминокислот из кишечника хозяина.
Тем не менее, аминокислота является важным источником энергии для вида. В основных аминокислотных путях используются аргинин и аспартат (Adam, 2000). Первый путь использует аргининдигидролазу и присутствует у многих прокариот, но на сегодняшний день только у двух эукариот.Этот путь превращает аргинин в аммиак и диоксид углерода и генерирует АТФ путем фосфорилирования на уровне субстрата. Орнитин, продукт, используется как мембранный антипорт для импорта внеклеточного аргинина (Edwards, 1992).
Импортные аминокислоты также используются для осморегуляции и защиты. Когда осмолярность окружающей среды падает, внутриклеточный аланин выводится из организма (Edwards et al, 1993 и Nygaard et al, 1994). Антипорт также включает транспорт L-серина, глицина, L-треонина, L-глутамина и L-аспарагина.Установлено, что цистеин играет роль в защите от кислородного отравления. Цистеин является основным источником свободных тиоловых групп на вариабельных поверхностных белках (VSP) трофозоитов, и это было показано с помощью радиоактивно меченного цистеина (Aggarwal et al, 1989).
Водородный обмен
G. lamblia , как обнаружено, генерирует небольшое количество водорода (2 нмоль / мин / 10 E 7 организмов) в строгих анаэробных условиях (Llyod et al. 2002). Никаких идентифицируемых гидрогеносом не обнаружено, хотя они часто встречаются у других производящих водород видов, таких как трихомонады).
Экология
Жизненный цикл G. lamblia состоит из двух стадий: цисты и трофозоитов. Киста обладает высокой устойчивостью к хлорированию, озонолизу и может выжить в холодной воде при температуре от 4 до 8 градусов по Цельсию (Али и др., 2003). Часто встречается в зараженной воде и фекалиях инфицированного человека. Форма трофозоита является вегетативной формой G. lamblia и обнаруживается в тонком кишечнике инфицированных людей. Он также может быть обнаружен в их кале.Трофозоит почти исключительно находится в кишечнике человека, его основной нише.
Приобретение и Excystation
Обнаружение лямблий происходит главным образом через кисту, фекально-оральным путем или, в непромышленных регионах, через загрязненную воду. Киста защищена от желудочного сока хозяина стенкой кисты. Исследования показали, что его прохождение через желудок хозяина создает кислую среду, которая способствует инициированию эксцистанции, превращения кисты в трофозоит.(Элмендорф и др., 2003 г. и Адам 2001 г.) Эксцистации также способствует протеаза поджелудочной железы и цистеиновая протеаза CP2. Кроме того, были собраны доказательства, чтобы показать, что эксцистанция может ингибироваться антителами к стенке кисты, антагонистами кальмодулина TFP и W7 (Bernal et al, 1998) и агглютинином зародышей пшеницы (Meng et al, 1996).
Приложение и размножение
Трофозоит прикрепляется к стенке тонкой кишки с помощью вентрального диска и размножается в просвете.Считалось, что прикрепление осуществляется за счет гидродинамической силы, создаваемой под диском с помощью вентральных жгутиков (Holberton et al, 1974). Другой предложенный механизм прикрепления включает поверхностный лектин, обнаруженный на вентральном диске трофозоитов, который связывается с маннозой. (Фартинг и др., 1986).
Размножение и инцистация (образование кисты) осуществляется в средней и средней части тощей кишки (тонкий кишечник) хозяина. Метаболическая активность трофозоитов может вызывать диарею, мальабсорбцию и потерю веса - состояние, известное как лямблиоз [подробнее см. Патология].У некоторых пациентов симптомы могут не развиваться. Причина этого неизвестна, но считается, что штамм паразита и факторы хозяина могут иметь значение (Thomson, 2000).
Энцистанция
Encystation можно наблюдать под микроскопом, а подробные механизмы описаны биохимическими методами.
Энцистация происходит при слабом щелочном pH 7,8 в присутствии жирных кислот, конъюгированных с солями желчных кислот (Gillin et al, 1988). Исследования in vitro показали, что инцистация вызывается холестерином и воздействием солей желчных кислот и жирных кислот (Kane et al, 1991 и Gillin et al, 1996), но это остается спорным.
Есть две фазы энцистации. На ранней стадии белки клеточной стенки синтезируются и транспортируются на периферию. Повышенный синтез белков клеточной стенки делает структуру, подобную Гольджи, легко видимой с помощью электронной микроскопии (Reiner et al, 1990). Белки, содержащие сигнатуру, богатую лейцином, перенаправляются на специфичные для энцистации транспортные везикулы (ESV), видимые под световой микроскопией (Hehl et al, 2000).
Поздняя фаза включает сборку стенки кисты и морфологические изменения.Сборка начинается с филаментов стенки кисты, затем с филаментного слоя клеточной стенки (Adam, 2001). В ходе поздней фазы энцистации трофозоит теряет подвижность. В итоге получается неприлипающая киста, включающая два физически соединенных трофозоита с четырьмя ядрами.
Патология
Лямблиоз является клиническим проявлением инфекции G. lamblia . Он характеризуется сильной диареей, недостаточным питанием, потерей веса и легкими повреждениями эпителия кишечника.Полный патогенез неизвестен, но существует несколько теорий.
Предполагается, что нарушение эпителия тонкой кишки вызвано индуцированным апоптозом (Chin et al, 2002). Недавно эта теория была доказана in vitro. Наблюдались как внутренние, так и внешние пути апоптоза энтероцитов. Кроме того, Bcl-2 подавлялся, и было проанализировано действие каспазы-3 (Panaro et al, 2007). Однако эта теория кажется специфичной для штамма. Также было показано, что G.lamblia увеличивает проницаемость однослойного эпителия двенадцатиперстной кишки in vitro за счет фосфорилирования F-актина и зон окклюзии в энтероцитах (Scott et al, 2002).
Кроме того, иммунное уклонение G. lamblia достигается за счет вариации экспрессии вариантно-специфических поверхностных белков (VSP), семейства вариабельных антигенных создателей неизвестной функции (Singer et al, 2000). Селективное давление на паразита бывает как положительным, так и отрицательным и зависит от ВСП. Также могут быть задействованы специфические для хозяина факторы.
Хотя наиболее яркими клиническими проявлениями лямблиоза являются диарея и мальабсорбция, подробные механизмы не описаны полностью. Однако колонизация микробом, по-видимому, вызывает укорочение микроворсинок (Scott et al, 2000), уплощение ворсинок или атрофию (Williamson et al., 2000). Считается, что эти механизмы могут работать синергетически с другими механизмами, такими как включение дисахаразы и протеазы, вызывая общие симптомы (Muller et al, 2005).
Недавно проведенное исследование показало, что при лямблиозе диарея является причинной комбинацией утечки, мальабсорбции и мальсекреции: подавление протеина плотного соединения клаудина 1 и эпителиального апотоза вызывает нарушение натрий-зависимой абсорбции глюкозы, что приводит к активному хлориду. секреция ионов. Следовательно, вода попадает в просвет, вызывая диарею (Troeger, 2007).
Применение в биотехнологии
На данный момент не было задокументировано.
Текущие исследования
г.lamblia тщательно изучается из-за его значения в эпидемиологии, клинической медицине и эволюционной биологии.
- В связи с широким распространением болезней, передаваемых через воду, вызываемых G. lamblia , это одно из основных направлений исследований эпидемиологов. Poereira и др. Оценили распространенность лямблиоза среди госпитализированных детей, страдающих диареей, в Гоянии, Бразилия, и определили несколько факторов риска инфекции G. lamblia (Pereira et al, 2007).Эти факторы риска включают: количество детей в семье, гигиену питания, присутствие кошек и посещение детских садов. Подобные исследования в настоящее время проводятся CDC и другими правительственными агентствами по всему миру (CDC, 2004). Выявление таких факторов риска позволяет лицам, определяющим политику, бороться с болезнью с точки зрения законодательства и служб общественного здравоохранения.
- Изучена лекарственная устойчивость G. lamblia (клоны штамма WB C6) к нитазоксаниду и метронидазолу (Muller et al, 2007).Транскрипция нескольких белков, участвующих в формировании устойчивости, таких как пируватоксиредуктазы 1 и 2, нитроредуктаза, VSP и протеин-дисульфид-изомеразы 2 и 4, была количественно оценена и коррелирована с каждым штаммом. И было обнаружено, что у каждого штамма синтез определенного набора белков подавлялся. Более того, было обнаружено, что экспрессия VSP резко изменилась под действием препаратов. Те же исследователи планируют объединить геномные и протеономические методы, чтобы детально понять клеточные механизмы и их роль в устойчивости паразита к лекарствам.
- Изучение генетической структуры G. lamblia (Yee et al, 2007) выявило гены ядерных гистонов h3a, h3b, h4 и h5, но не ген линкерного гистона h2. Сравнение последовательностей показывает сходные последовательности как в кодирующей, так и в 5’-некодирующей областях. Это свидетельствует о недавней дупликации или конверсии гена. Это исследование предлагает альтернативный механизм уплотнения генов в G. lamblia , который может быть осуществлен без гистона h2, чтобы оптимизировать транскрипцию богатой генами ДНК этого вида.Такое понимание было бы основополагающим в знании экспрессии генов у этого вида.
- Проект генома Giardia lamblia в Морской биологической лаборатории в Masshuttes, в котором секвенирование дробовика используется для получения полного генома G. lamblia , в настоящее время находится в стадии реализации. Аннотированные сборки генома выпускаются регулярно и доступны исследователям во всем мире. Анализ функции генов в настоящее время проводится в сотрудничестве с доктором Фрэнсис Гиллин из отдела молекулярной патологии Калифорнийского университета в США.Это исследование имеет большое значение для эволюционной биологии, так как оно даст представление об истории G. lamblia ': его связи с митохондриями и его роли в качестве моста между прокариотами и эукариотами. Полный геном также поможет исследователям идентифицировать возможные гены, кодирующие токсины, и, возможно, получить лекарства, которые могут подавить паразита.
Ссылки
- Адам, Р. Д. и Адам. 2001. Биология лямблий.Обзоры клинической микробиологии, 14 (3), 447-.
- Aggarwal, A., J. W. Merritt, Jr., and T. E. Nash. 1989. Богатые цистеином варианты поверхностных белков Giardia lamblia. Мол. Biochem. Паразитол. 32: 39–47.
- Али, С.А., и Хилл, Д.Р. 2003. Лямблии кишечные. Текущее мнение в области инфекционных болезней, 16 (5), 453-60.
- Бернал Р. М., Р. Товар, Дж. И. Сантос и М. Л. Муньос. 1998. Возможная роль кальмодулина в эксцистации Giardia lamblia. Паразитол. Res. 84: 687–693.
- Браун, Д.М., Апкрофт, Дж. А., Апкрофт, П. 1995. Свободнорадикальная детоксикация при Giardia duodenalis. Mol Biochem Parasitol 72: 47–56.
- Браун, Д.М., Апкрофт, Дж. А., Апкрофт, П. 1996b. Класс тиоредоксинредуктазы дисульфидредуктазы у простейших паразитов Giardia duodenalis. Mol Biochem Parasitol 83: 211–220.
- Центр контроля заболеваний, Информационный бюллетень по инфекции лямблиозами. 2004. Проверено 29 августа 2007.
- Chin AC, Teoh DA, Scott KG, et al. 2002. Штамм-зависимая индукция апоптоза энтероцитов с помощью Giardia lamblia нарушает функцию эпителиального барьера каспазо-3-зависимым образом.Infect Immun 2002; 70: 3673–3680.
- Corrêa, G, Morgado-Diaz, JA, & Benchimol, M., 2004. Центрин в Giardia lamblia - ультраструктурная локализация. Письма FEMS по микробиологии, 233 (1), 91-6.
- Эдвардс, М. Р., Л. А. Кнодлер, Дж. Р. Уилсон и П. Дж. Шофилд. 1993. Транспорт и метаболизм аланина Giardia Кишечник. Мол. Biochem. Паразитол. 61: 49–57.
- Эдвардс, М. Р., П. Дж. Шофилд, У. Дж. О’Салливан и М. Костелло. 1992. Метаболизм аргинина при культивировании Giardia Кишечник.Мол. Biochem. Паразитол. 53: 97–103.
- Элмендорф, Х.Г., Доусон, Южная Каролина, и Маккаффери, Дж. М.. 2003. Цитоскелет Giardia lamblia. Международный журнал паразитологии, 33 (1), 3-28.
- Фартинг MJG, Pereira MEA, Keusch GT 1986. Описание и характеристика поверхностного лектина из Giardia lamblia. Infect Immunol 1986; 51: 661-7.
- Фили, Д. Э., Дж. В. Шоллмейер и С. Л. Эрландсен. 1982. Giardia spp .: Распределение сократительных белков в прикрепительной органелле.Exp. Паразитол. 53: 145–154.
- Гиллин, Ф. Д., Д. С. Райнер и С. Э. Баучер. 1988. Факторы тонкого кишечника способствуют инцистации Giardia lamblia in vitro. Заразить. Иммун. 56: 705–707.
- Гиллин, Ф. Д., Райнер, Д. С., и Маккаффери, Дж. М. 1996. Клеточная биология примитивных эукариот Giardia lamblia. Анну. Rev. Microbiol. 50, 679-705
- Хашимото Т., Санчес Л. Б., Ширакура Т., Мюллер М., Хасегава М. 1998. Вторичное отсутствие митохондрий в Giardia lamblia и Trichomonas vaginalis, выявленное филогенией валил-тРНК синтетазы.Proc Natl Acad Sci USA 1998; 95: 6860-5.
- Хашимото Т., Л. Б. Санчес, Т. Ширакура, М. Мюллер и М. Хасегава. 1998. Вторичное отсутствие митохондрий у Giardia lamblia и Trichomonas vaginalis, выявленное филогенией валил-тРНК синтетазы. Proc. Natl. Акад. Sci. США 95: 6860–6865.
- Hehl, A. B., M. Marti, and P. Kohler. 2000. Стадия-специфическая экспрессия и нацеливание химер белок стенки кисты-зеленый флуоресцентный белок у лямблий. Мол. Биол. Ячейка 11: 1789–1800.
- Холбертон Д.В. и Маршалл Дж.1995. Анализ консенсусных последовательностей в промоторах гена цитоскелета лямблий. Nucleic Acids Res 1995; 23: 2945-53. Ссылка.
- Холбертон Д.В. 1974. Приложение Giardia - гидродинамическая модель, основанная на активности жгутиков. Журнал Эксперта Биол 1974; 60: 207-21.
- Кейн, А. В., Уорд, Х. Д., Кеуш, Г. Т. и Перейра, М. Е. А. 1991. Инцистирование Giardia lamblia in vitro: крупномасштабное производство цист in vitro и различия штаммов и клонов в эффективности энцистации. J. Parasitol. 77, 974-981
- Ladeira, RB, Freitas, MA, Silva, EF, et al.2005. Гликоген как углеводный запас энергии в трофозоитах Giardia lamblia. Паразитологические исследования, 96 (6), 418-21.
- Ле Блан, С. М., Р. С. Касе, и Л. Х. Ван дер Плоег. 1991. Анализ рРНК Giardia lamblia, кодирующей теломер с [TAGGG] n в качестве теломерного повтора. Nucleic Acids Res. 19: 5790–5790.
- Lei Li, Ching C. Wang 2006. Вероятная молекулярная основа восприимчивости лямблий к кислороду Молекулярная микробиология 59 (1), 202–211.
- Ллойд, Д., Харрис, Дж. К., Марулис, С. и др.2002. «Примитивный» микроаэрофил Giardia Кишечник (син. Lamblia, duodenalis) имеет специализированные мембраны с функциями транспорта электронов и генерации мембранного потенциала. Microbiology, 148 (Pt 5), 1349-54.
- Лухан, Х. Д., А. Маротта, М. Р. Моватт, Н. Шаки, Дж. Липпинкотт-Шварц и Т. Э. Нэш. 1995. Индукция развития структуры и функции Гольджи у примитивных эукариот Giardia lamblia. J. Biol. Chem. 270: 4612–4618.
- Мэн Т.С., М.Л. Хецко и Ф.Д. Гиллин. 1996. Ингибирование эксцистации Giardia lamblia антителами против стенок кисты и агглютинином зародышей пшеницы. Заразить. Иммун. 64: 2151–2157.
- Michels, PA, & Rigden, DJ. 2006. Эволюционный анализ метаболизма 2,6-бисфосфата фруктозы. ИУБМБ жизнь, 58 (3), 133-41.
- Muller N, Allmen NV. 2005. Недавние исследования реакций слизистых оболочек, связанных с инфекциями Giardia lamblia. Международный журнал паразитологии 35 (2005) 1339–1347
- Muller, J, & Muller.2007. Характеристика клонов Giardia lamblia WB C6, устойчивых к нитазоксаниду и метронидазолу. Журнал антимикробной химиотерапии, 60 (2), 280-.
- Найгаард Т., К. К. Беннет, Г. Гроссман, М. Р. Эдвардс и П. Дж. Шофилд. 1994. Отток аланина Giardia Кишечник. Мол. Biochem. Паразитол. 64: 145–152.
- Пэджет, Т. А., Э. Л. Джарролл, П. Мэннинг, Д. Г. Линдмарк и Д. Ллойд. 1989. Дыхание в цистах и трофозоитах Giardia muris. J. Gen. Microbiol.135: 145–154.
- Panaro MA, Cianciulli A, Mitolo V, Mitolo C.I, Acquafredda A, Brandonisio O, Cavallo P. 2007. Каспазозависимый апоптоз эпителиальной клеточной линии HCT-8, индуцированный паразитом Giardia Кишечник. FEMS Immunol Med Microbiol. 2007 Aug 20;
- Papanastasiou, P., T. Bruderer, Y. Li, C. Bommeli, and P. Kohler. 1997. Первичная структура и биохимические свойства вариантно-специфического поверхностного белка лямблий. Мол. Biochem. Паразитол. 86: 13–27.
- Pereira, MGC, ATWILL, ER и BARBOSA, AP.2007. Распространенность и связанные факторы риска заражения Giardia lamblia среди детей, госпитализированных с диареей в Гоянии, штат Гояс, Бразилия. Rev. Inst. Med. троп. С. Пауло, май / июнь 2007 г., том 49, номер 3, стр.139-145
- Райнер Д. С., М. Маккаффери и Ф. Д. Гиллин. 1990. Сортировка белков стенки кисты по регулируемому секреторному пути во время дифференцировки примитивного эукариота, Giardia lamblia. Евро. J. Cell Biol. 53: 142–153.
- Roger AJ, Svard SG, Tovar J et al. 1998 г.Митохондриально-подобный ген шаперонина 60 у Giardia lamblia: свидетельство того, что на дипломатонаде когда-то был эндосимбионт, родственный предку митохондрий. 1998. Proc Natl Acad Sci USA, 95: 229-34.
- Санчес, Л. Б., Элмендорф, Х., Нэш, Т. Е., Мюллер, М., 2001. NAD (P) H: менадион-оксидоредуктаза амитохондриального эукариота Giardia lamblia: более простой гомолог фермента позвоночных. Микробиология 147: 561–570.
- Scott KG, Meddings JB, Kirk DR, et al. 2002. Кишечная инфекция, вызванная Giardia spp.снижает функцию эпителиального барьера в зависимости от киназы легкой цепи миозина. Гастроэнтерология 123: 1179–1190.
- Скотт, К.Г., Логан, М.Р., Кламмер, Г.М., Теох, Д.А., Бурет, А.Г., 2000. Микроворсинки щеточной границы Джеюнала у мышей, инфицированных Giardia muris: роль Т-лимфоцитов и интерлейкина-6. Заразить. Иммун. 68, 3412–3418.
- Зингер С.М., Элмендорф Х.Г., Конрад Дж. Т., Нэш Т.Э. 2000. Биологический отбор вариантно-специфических поверхностных белков в Giardia lamblia. J. Infectious Diseases 2001; 183: 119-124 .
- Солтыс, Б.Дж., М. Фалах и Р. С. Гупта. 1996. Идентификация эндоплазматического ретикулума у примитивных эукариот Giardia lamblia с использованием криоэлектронной микроскопии и антител к Bip. J. Cell Sci. 109: 1909–1917.
- Теодорович, С. Уоллс, CD. и Элмендорф, HG. 2007. Двунаправленная транскрипция является неотъемлемой особенностью промоторов Giardia lamblia и вносит вклад в обилие стерильных антисмысловых транскриптов по всему геному. Исследование нуклеиновых кислот, 2007, Vol. 35, № 8 2544-2553. Ссылка.
- Томпсон, Р.C.A., 2000. Лямблиоз как повторно возникающее инфекционное заболевание и его зоонозный потенциал. Int. J. Parasitol. 30. С. 1259–1267.
- Troeger, H, Epple, HJ, Schneider, T. и др., 2007. Влияние хронической инфекции Giardia lamblia на транспорт эпителия и барьерную функцию в двенадцатиперстной кишке человека. Гут, 56 (3), 328-35.
- Yee, J, Tang, A, Lau, WL, et al. 2007. Основные гистоновые гены Giardia Кишечник: геномная организация, структура промотора и экспрессия. BMC молекулярная биология, 8, 26-.
- Ю., ЛЗ. Бирки, К. Виллиа-младший, Адам, Род. 2002. Каждое из двух ядер лямблий имеет полные копии генома и равномерно делится при цитокинезе. ЭУКАРИОТИЧЕСКАЯ КЛЕТКА, апрель 2002 г., стр. 191–199 a
Отредактировал студент Д. Лау из Рэйчел Ларсен
Под редакцией KLB
.лямблий и домашних животных | Лямблии | Паразиты
Giardia трудно полностью исключить из окружающей среды, но есть вещи, которые вы можете сделать, чтобы снизить риск повторного заражения домашних животных и заражения человека. Помните, что, несмотря на все ваши усилия по очистке окружающей среды, Giardia может сохраняться на открытом воздухе, и возможно повторное заражение домашних животных 4 .
- При работе с фекалиями надевайте перчатки.
- Немедленно удалите кал 4 и поместите в пластиковый пакет.
- По возможности ограничьте доступ к общим открытым пространствам, если домашние животные страдают диареей или проходят лечение от Giardia .
- Устраните любые источники стоячей воды (например, лужи, емкости с водой и неиспользуемые фонтаны).
- Не пытайтесь использовать отбеливатель или QATS для обработки почвы или травы, так как они будут неэффективными.
- Не допускайте позволять новым животным, особенно молодым, заходить во двор или другое открытое пространство до получения разрешения ветеринара.
Giardia lamblia | протист | Britannica
Giardia lamblia , также называемая G. Кишечник , одноклеточный паразит отряда Diplomonadida. Как и у других дипломонад, клетки G. lamblia имеют два ядра и восемь жгутиков. Паразит прикрепляется к слизистой оболочке кишечника человека с помощью сосательного органа, вызывая диахреальное состояние, известное как лямблиоз. Острый лямблиоз - распространенное заболевание среди туристов, отдыхающих и путешественников в неразвитые страны, которые пьют неочищенную воду, а также среди детей в детских садах и людей, пользующихся переполненными местами для купания.Клинические симптомы включают обильный жирный стул с неприятным запахом, спазмы желудка и вздутие живота. Паразит передается непосредственно со стулом инфицированных людей или животных лицам, употребляющим зараженную воду, пищу или другие материалы. Симптомы появляются после 10-дневного инкубационного периода и могут сохраняться в течение нескольких недель. Заболевание обычно протекает в легкой форме и проходит самостоятельно, хотя у инфицированных детей может развиться хроническое нарушение всасывания питательных веществ из кишечника. Его лечат антимикробными препаратами и жидкостями для предотвращения обезвоживания.
Giardia lamblia
J.M. Langham .